进化介绍人类进化英文中的“”一词
进化的定义是什么?
进化,又称演化(),在生物学中是指种群里的遗传性状在世代之间的变化。所谓性状是指基因的表现,在繁殖过程中,基因会经复制并传递到子代,基因的突变可使性状改变,进而造成个体之间的遗传变异。 新性状又会因物种迁徙或是物种间的水平基因转移,而随着基因在种群中传递。当这些遗传变异受到非随机的自然选择或随机的遗传漂变影响,在种群中变得较为普遍或不再稀有时,就表示发生了进化。简略地说,进化的实质便是:种群基因频率的改变。 进化进化介绍人类进化英文中的“”一词,起源于拉丁文的“”,原意是将一个卷在一起的东西打开,也指任何事物的生长、变化或发展,包括恒...全部
进化,又称演化(),在生物学中是指种群里的遗传性状在世代之间的变化。所谓性状是指基因的表现,在繁殖过程中,基因会经复制并传递到子代,基因的突变可使性状改变,进而造成个体之间的遗传变异。
新性状又会因物种迁徙或是物种间的水平基因转移,而随着基因在种群中传递。当这些遗传变异受到非随机的自然选择或随机的遗传漂变影响,在种群中变得较为普遍或不再稀有时,就表示发生了进化。简略地说,进化的实质便是:种群基因频率的改变。
进化进化介绍人类进化英文中的“”一词,起源于拉丁文的“”,原意是将一个卷在一起的东西打开,也指任何事物的生长、变化或发展,包括恒星的演变,化学的演变,文化的演变或者观念的演变。
自从19世纪以后,演化通常用来指生物学上,不同世代之间外表特征与基因频率的改变。达尔文并未对“”下过定义,在1859年出版的《物种起源》第一版中,也并未使用“”这个字,当时达尔文是使用“经过改变的继承”( with )、“改变过程”( )或是“物种改变的原理”( of the of )等词。
这个字在当时生物学上的意义,是胚胎发育过程,在当时一般用语中具有“进步”的含义,而达尔文反对将“进步”之类的用语来描述生物改变的过程。他曾在《物种起源》第7章中说:“(天择)的最后结果,包括了生物体的进步()及退步()两种现象”。
而后来包括达尔文在内的学者,之所以改用来描述生物演化现象,乃因英国哲学家赫伯特·史宾赛在许多著作里进行的名词统一。以自然选择为基础的进化理论,最早是由查尔斯·达尔文与亚尔佛德·罗素·华莱士所提出,详细论述则出现在达尔文出版于1859年的《物种起源》。
20世纪30年代,达尔文自然选择说与孟德尔遗传合二为一,形成了现代综合进化理论,连结了进化的“机制”(自然选择)与进化的“单位”(基因)。这种有力的解释及具预测性的理论,成为现代生物学的中心原则化进化退,使地球生物的多样性得以统一说明。
转座子的坟场——人类基因组进化主要学说(1)拉马克的获得性状遗传学说(2)达尔文的自然选择学说(3)直生论(4)生物进化的“突变论”(5)新达尔文主义学说(6)综合进化学说(7)中性学说(8)“间断平衡”学说(9)进化“四因说”进化进化与演化进化概况最初中文对“”这个字有两种翻译。
“进化”一词来自日语(日制汉语)。严复是最早反对使用“进化”者之一。后人在《天演论》书尾的名词表中写到:“一词,严氏译为天演,近人撰述多以进化二字当之。严复主张以'天演'取代'进化'。
”《天演论》中文对于如何翻译“”颇有争议。支持使用“演化”的学者认为,演化在字面上的意义比较中性,能表达连续与随机的意义,进化则带有“进步”的含意。由于汉语中“进”与“退”是代表相反意义的两个字,因此若使用进化,则在逻辑上不易将“退化”定义为进化的一种类型。
对翻译的争论也表现了人们对进化论理解的变化,过去“进化”多表示生物朝适应环境的方向演化,而当时许多人认为生物的演化是随机的,并没有进步退步之分。严复所翻译的不是《物种起源》,而是赫胥黎的《进化论与伦理》,多部分内容与生物学无关。
赫胥黎于该书导言中设有一节,立之界说,“谓为初指进化而言,继则兼包退化之义。严氏于此节略而未译,然其用天演两字,固守赫氏之说也”。也就是说,严复理解到单用“进化”一词的缺憾,故自创“天演”二字取代“进化”。
早在达尔文逝世不久时,中文上用“进化论”之名是一致的,例如孙中山在著作中盛赞该论。除了台湾,“进化论”一词沿用至今。台湾的学者、政府机构、教科书等也一向沿用“进化论”,台湾教育部却更改课本,以演化论代替,新旧学生被概念混淆,不知其是否为同样理论或是两套理论。
根据台湾教育部所编辑的辞典,“进化”定义为生物由低级到高级、由简单到复杂的发展过程,并将“退化”定义为进化的反义词。演化则被定义为生物物种为了因应时空的嬗变,在形态和行为上与远祖有所差异的现象,演化明显较适用于生物学的讨论。
在翻译来说,主要考量的是达尔文所取英语原名的意义,由简单到复杂生物的出现确是有“进”的含义。而与达尔文自然选择论相对抗的如“智慧设计”论等,则否认眼睛这样精确和复杂的器官可以进化出来。进化演化模型微观演化与宏观演化演化可以依据时间长短与差异程度,分成“微观演化”(微演化)与“宏观演化”(广演化)。
微观演化指几个世代中,基因频率小范围的变化,例如现今世界各地人类的差异。宏观演化指长时间的演化过程,例如人类与灭绝祖先的关系。宏观演化的历史,包括生物群在化石纪录中突然出现突然丢失的环节、物种长期停滞不变等难以解释的现象,例如延续时间5370万年左右的寒武纪地层中保存大量化石的现象。
演化速率红原鸡的演变关于宏观演化的过程,有许多不同的理论,主要是用来解释化石纪录中难以解释的现象。传统的观念认为,许多微观演化的累积,经历足够时间之后便形成新物种,这样的理论称为渐变论。而渐变论又有许多形式,例如由史蒂芬·古尔德与尼尔斯·艾崔奇在1972年提出的疾变平衡论。
这种理论认为生物的演化历史是许多走走停停的过程,在大多数时间并没有太大的变化。而某些个体的关键基因(如同源异形盒),可能导致新物种迅速形成并大量繁殖,之后再恢复平衡。如此便能够合理解释化石纪录不平均的问题。
古尔德经常以批判渐变论的姿态出现在大众媒体上,而道金斯认为进化介绍人类进化英文中的“”一词,疾变平衡论只是渐变论的一种形式。极端的渐变论认为,演化过程是以等速进行的,对于化石数量不平均问题,以“化石纪录本身并不完整”来解释。
极端的渐变论,至今仍然经常使用在分子层次,并发展出分子时钟的观念。研究发现,分子时钟也并非等速变化,而是在不同物种或是不同年代具有不同的演化速度。不同的研究中,对分子演化速率的推算差异相当大。
另外有一种跃进论,认为物种只需要一个世代就能够形成,且可能具有创造论与灾变论等形式。创造论与灾变论都是源自宗教,如《创世纪》与大洪水,不过灾变论将火山爆发、陨石撞击等所有来自地球内外的影响包含在内。
进化数学模式生物的变异可以量化为数值,因此也能够建立出数学模型。较早的数值分类学( )将生物的特征量化为数字,并且依照这些数字进行分类,借此找出它们的亲缘关系。
科学家一般认为生物型体的演化,源自于基因变异(基因中心演化观点)。由于基因突变具有一些规律性,因此复杂的演化过程,可以简化成数学模型。早期的新达尔文主义科学家使用线性的模型,例如将一个对偶基因定下一个“天择系数”,借此估计该对偶基因在每一世代中的基因频率。
现今则多用非线性的方式分析。例如一种称为生殖成就空间( )的曲面图形,可以用来表达生物个体在繁殖上的能力与其特征之间的相关性,由图形可以看出具有不同特征的生物可能同时具有不同的生殖成就。
由于这些生物的变异可以化成多种不同的变量,因此数学上处理多维空间的技巧,便可应用于生物关系的分析。此外数学方法的优点是计算过程相当精确,缺点是其正确性取决于人为的判断与假设。进化进化证据古生物学与化石纪录古生物学是以生物化石为基础,以研究生物亲缘关系的一种研究。
当生物个体死亡之后,它的尸体通常经由微生物分解而腐化,使得生存痕迹消失。但有时候这些遗迹可能会因为某些因素而被保存。只要是来自古代生物造成的痕迹,或是生物体本身,都可以称为化石。化石对于了解生物演化历程而言相当重要,因为它是较为直接的证据,且带有许多详细的资讯。
在化石形成过程中,生物体的痕迹由于快速地受到掩埋,因此不会发生风化与分解的情形。较为常见的化石,通常源自骨骼或外壳等坚硬部位,并经由类似铸模的过程形成。坚硬的骨骼在动物死亡之后,会因为有机物的腐败,而产生一些漏洞。
将骨骼掩埋的砂石或矿物,则会经由这些漏洞侵入骨骼内部,并将其填满。这种过程称为置换作用,属于型体的保留,而不是生物体本身的保留。也有一些化石是生物体本身,例如被冰冻的猛犸象、琥珀里的昆虫。此外,古代动物的脚印、或植物在地底下因温度与压力作用而碳化,都可称为化石。
澄江生物群不同时代的生物化石,会出现的在不同的地层中,如此便能够研究古生物之间,以及它们与现代生物之间的关系。“失落的环结”指演化过程可能出现过,却尚未发现的物种。连接两个物种之间的化石,则称为“过渡化石”。
例如可能位于鸟类与恐龙中间的始祖鸟()化石,以及一种具有四肢的大型浅水鱼()可能是鱼类与两栖类的过渡化石。化石纪录对于古生物的研究有所限制,因为形成化石并不容易。
举例而言,软体动物身上并没有太多能够形成化石的部分,还有一些生物生存在难以形成化石的环境当中。即使化石形成之后,也有可能因为某些原因被摧毁,使得大多数化石皆是零散的状态,只有少数化石能够保持完整。

而当演化上的改变在族群中只占有少部分,或是环境变化使族群规模缩小时,都会使它们形成化石的机率相对较小。此外,化石几乎无法用来研究生物内部器官构造和机制。寒武纪大爆发是达尔文以来困惑科学界一个多世纪的科学难题。
澄江动物群代表包括脊索动物在内的许多现生动物门和亚门的重要历史起点,为揭示寒武纪大爆发的性质和特征,见证高级分类单位起源在先、低级分类单位起源在后的“自上而下”的宏观演化模型,以及为探索动物(特别是脊索动物和节肢动物)起源等提供了重要证据,充实了演化论内涵。
最古老脊索动物化石的发现与研究首次再现了包括人类在内的脊椎动物的生命蓝本。这些化石(特别是海口虫)保存的完整性令人叹为观止,包括了脑、神经索、血管等软组织构造,既包含有脊椎动物特征,又具有相似文昌鱼的特征,为无脊椎动物与脊椎动物间建起了桥梁。
生物地理学与物种分布A图显示古代大陆间有陆桥连结,因此动物可在大陆间自由迁徙。B图显示陆桥消失之后,各地物种受到孤立化进化退,并形成新物种。生物地理(左A图右B图)由于板块移动造成的大陆漂移(如南美洲与非洲),以及冰河时期前后造成的海平面高度变化(如白令海峡陆桥),改变了陆地间的相连性。
一些相距遥远的地区,可在地下挖出许多相似的生物化石,而海洋或山脉的隔离作用,却使现有的物种具有相当大的差异。例如在南美洲的猴、美洲豹、骆马化进化退,与非洲的猴、狮子、长颈鹿。此外,与世界上其它地方的胎盘动物相比较为原始的有袋类动物,虽已大多灭绝,但澳大利亚大陆却依然保存着袋鼠无尾熊等许多有袋类动物。
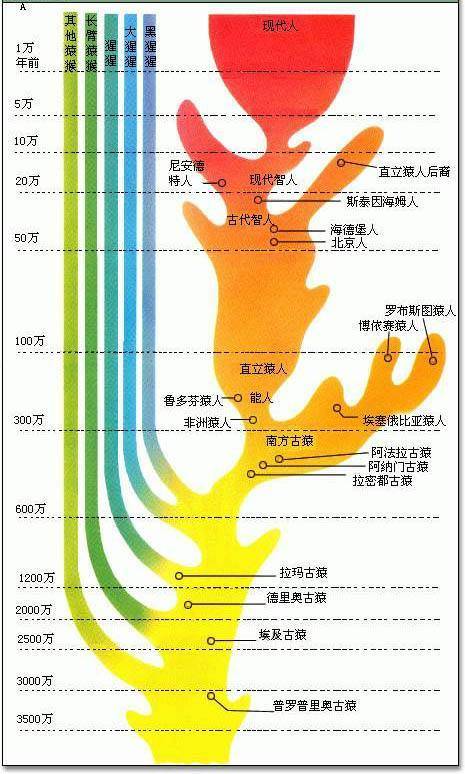
除不同大陆之间有这种现象,大陆与其邻近岛屿因曾在地理上相连,因而也能够找到相似但变异了的物种。例如台湾、中国大陆与日本的猕猴之间的差异。型态比较对脊椎动物五趾肢的比较,支持了脊椎动物具有共同祖先的理论。
举例而言,虽然人类、猫、鲸鱼与蝙蝠的五趾肢在型态上有所差异,但是主要架构都很相似。这些“同源”的构造,适应了不同的功能,如抓握、行走、游泳与飞行。此外有一些构造在功能上相似,但却具有不同的型态。
例如蝙蝠、鸟类与昆虫的翅膀;昆虫与脊椎动物的腿;章鱼与脊椎动物的眼睛;鱼类、鲸鱼与龙虾的鳍等。这类“异源”的构造,适应了相同的功能,如飞行、行走、感光与游泳。发育过程在发育初期都非常相似,在发育的过程中,这样的相似会逐渐减少,最后形成各物种的型态。

举例而言,虽然各种成熟的脊椎动物差异很大,但是它们的胚胎型态在发育初期却非常相似,腮裂仍然出现在已经没有腮的爬虫类、鸟类与哺乳类胚胎中。鱼类的二房心会被保留到成体,而人类的胚胎虽然也有这种构造,却会在胚胎成熟后消失。
胚胎19世纪70年代提出胚胎重演学说的恩斯特·海克尔,认为不同物种在同一段发育时期的差异也会显现出这些物种在演化上的亲近程度,而且这些生物演化历史的重复表现,能够出现在任何生物的胚胎发育过程。
但是到了1997年,海克尔的理论正式被推翻。理察森与先前的一些研究者发现,胚胎发育的型态,并不完全符合演化上的亲近程度。即使如此,现今的某些中学生物教科书依然继续使用海克尔的理论。在胚胎重演论提出直到被推翻的期间,胚胎学对于演化机制的解释并没有太大的进展。
但是演化发育生物学( ,)研究,将分子生物学与发育生物学等学科结合,解释基因的改变对于动物形态的控制过程。同时也发现外表差异相当大的动物之间,也拥有相同的调控基因。
以及相同的基因在不同的时间与空间,具有不同的作用。这些调控动物发育过程的基因,主要为一类Hox基因。分子证据分子生物学与生物化学可以利用构成生命的各种分子,来研究生物的亲缘关系。这些分子包括了核糖核酸(RNA)、脱氧核糖核酸(DNA)、蛋白质与糖体等。
科学家已经可以用机器与电脑将这些分子的碱基序列、氨基酸序列定序。研究这些序列的学科分别为基因体学、蛋白质体学。此外还有一种更为复杂的糖体学,专门分析糖类构造。基因体学透过脱氧核糖核酸序列的比较进化介绍人类进化英文中的“”一词,可以建立出与传统分类学大致符合的种系发生树。
这种方法通常用来确认并增强分类学的研究,将其它演化证据的漏洞修补或是修正。例如人类的DNA序列与黑猩猩的差异约为1。2%,与大猩猩的差异约为1。6%,与狒狒则为6。6%,如此就能推断并量化人类与猿类的亲缘关系。
此外一种组成核糖体的序列,也是研究亲缘关系的重要分子,这些研究导致了三域系统的出现,将真核生物、古生菌与细菌分开。蛋白质体学也支持了生物具有共同祖先的说法,因为许多生命所需的蛋白质,例如核糖体、DNA聚合酶与RNA聚合酶,不但出现在较原始的细菌中,也出在复杂的哺乳类体内。
这些蛋白质的核心部分在不同生物中保有相似的构造与功能,而较复杂的生物具有较多的蛋白质次单位( ),以调控更复杂的蛋白质相互作用。生物体内有一种脱氧核糖核酸片段,虽然与已知基因具有相似的序列,但是却无法发挥作用并完成蛋白质的制造。
这类无用的核酸片段称为伪基因,是支持生物具有共同祖先的证据之一。它们之所以无法正常作用,是因为在演化过程中,基因突变累积过多的缘故。抗药性微生物与病毒的抗药性,在医学上是重要问题,也是演化证据之一。
例如金黄葡萄球菌在1943年时仍可使用青霉素(盘尼西林)治疗,到了1947年就已经发现具抗药性的菌株。20世纪60年代改用甲氧苯青霉素,同样因为抗药性菌种的散布,使得20世纪80年代改用万古霉素,2002年时,已发现抗万古霉素的菌种。
标签: 进化 演化 生物 化石 基因六爻专家网